Conservation and Movement Ecology
I am an ecologist, interested in developing
methodology and theory to inform wildlife management decisions in a changing world
methodology and theory to inform wildlife management decisions in a changing world


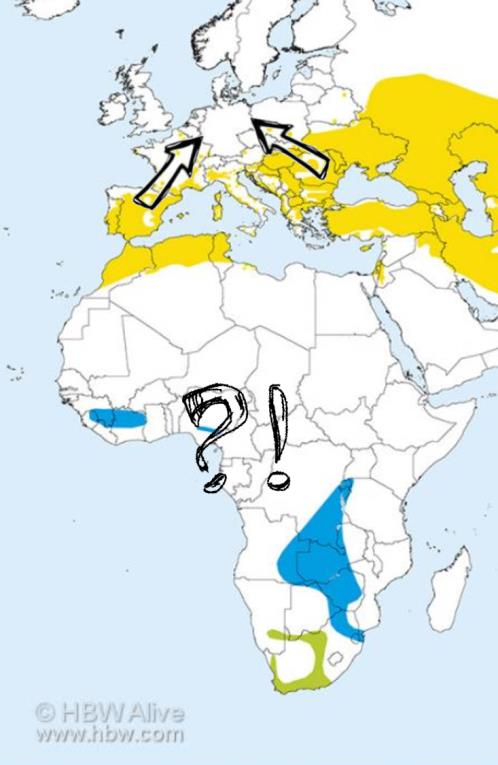







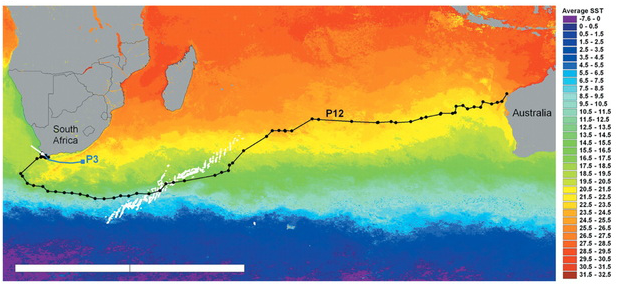

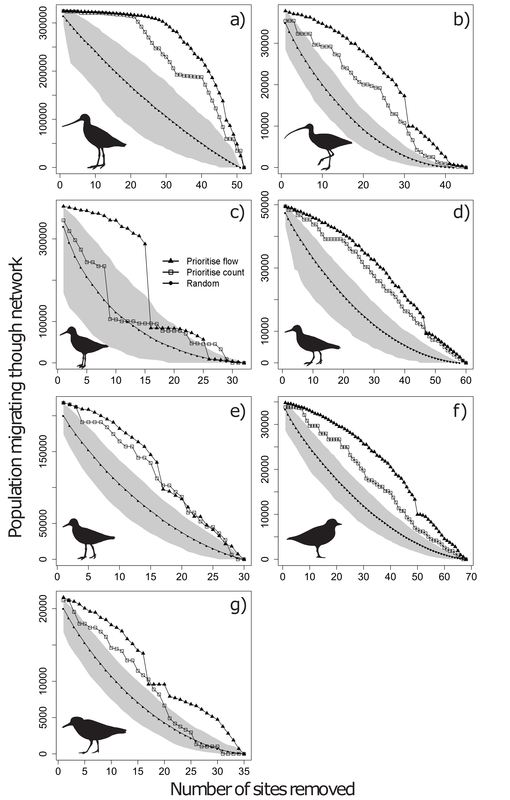

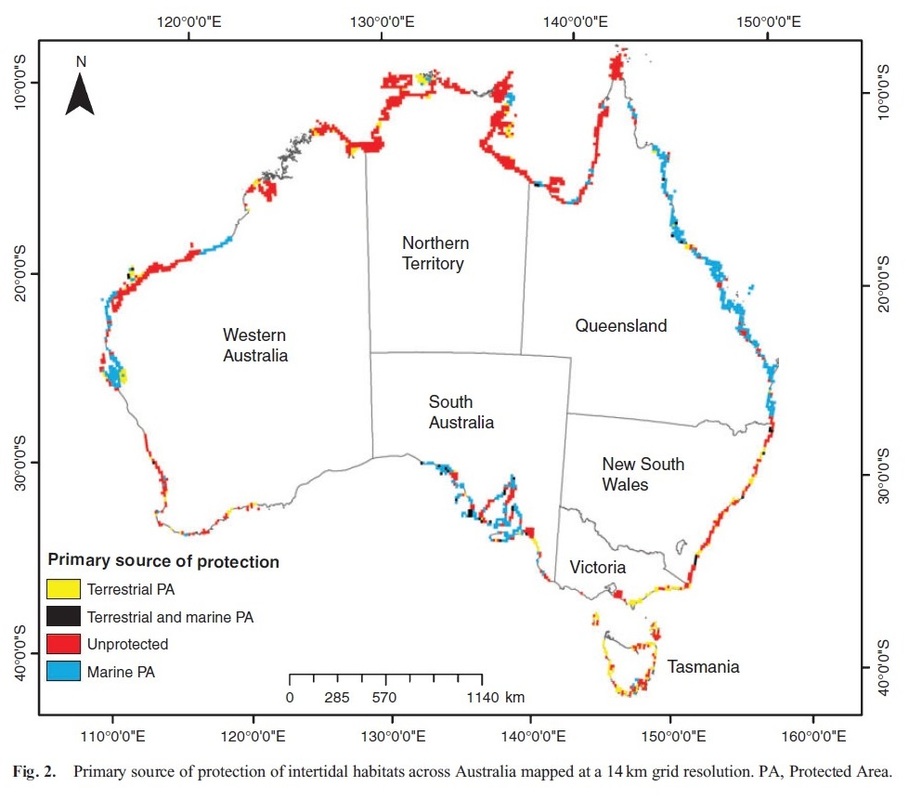




 RSS Feed
RSS Feed
